While not as persistent as gravity, plants regularly encounter touch stimuli in their environment. For twining plants, the presence of a solid support induces differential growth of the tendrils, resulting in the familiar coiling response and the association with the support. For roots, touch sensing takes the form of avoiding obstacles in the path of elongation, resulting in the redirection of the root around the object. For stems, touch sensing may mean the perception of wind and other forces that shake the stem, leading to its increased radial thickening and rigidity. Without this rigidification in response to mechanical stimuli, plants would probably collapse under their own weight.
Touch stimulation results in both rapid changes within cells and longer-lived changes in development. One of the first events observed after a touch stimulus is an increase in intracellular calcium ions (Ca2+). The release of Ca2+ is probably linked to the temporary mechanical deformation of the plasma membrane, which contains mechanosensitive ion channels. One popular model holds that these channels open upon a mechanical stimulus, thus allowing the influx of Ca2+ leading to the elevation of intracellular concentrations.
The elevated Ca2+ concentration within the cell induces two other rapid changes in response to touch. High Ca2+ inhibits the plasma membrane H+-ATPase and probably also stimulates the opening of inward rectifying H+ channels, both of which drive an increase in intracellular H+ levels and an increasing alkalinization of the apoplast. Elevated Ca2+ also stimulates the production of reactive oxygen species outside the cell by activating an enzyme in the plasma membrane. Both ROS and alkalinization of the apoplast are likely associated with increasing the rigidity of the cell wall as a result of mechanical stimulation. Interestingly, both the acidification of the cytoplasm and ROS production can be induced by artificially increasing Ca2+ levels.
Longer-term responses to touch involve the synthesis of the gaseous plant hormone ethylene, as well as the upregulation of a variety of genes, including the TCH genes. Several of the TCH genes encode Ca2+ binding proteins such as calmodulins or proteins like calmodulins, which bind Ca2+ and mediate downstream responses. Another of the TCH genes (TCH4) encodes a xyloglucan endotransglycosylase, an enzyme involved in modifying cell wall polysaccharides.
References
Monshausen GB, Bibikova TN, Weisenseel MH, Gilroy S (2009). Ca2+ regulates reactive oxygen species production and pH during mechanosensing in Arabidopsis roots. The Plant Cell 21: 2341–2356
Plants respond to unilateral light by growing toward the light source (in the case of shoots) or away from the light source (in the case of roots). This response, known as phototropism, has an action spectrum with a broad peak in the blue region, and a maximum at approximately 450 nm. A biochemical approach isolated several membrane-bound proteins whose phosphorylation was required for the phototropic response, implicating a common signal transduction component in the plant’s response to unilateral light.
Using a mutant screen in the model plant Arabidopsis, several novel mutants were isolated that showed non-phototropic hypocotyls (the nph mutants). Cloning and characterization of the nph1 mutant revealed it to encode a polypeptide with several intriguing domains. On one end of the protein was a kinase domain, responsible for phosphorylating a target substrate, and on the other end of the protein was a LOV domain. LOV, (for light, oxygen, and voltage sensing), is an evolutionarily conserved domain found in a number of organisms and involved in sensing one of these physical cues. The protein encoded by NPH1, which has been named PHOTOTROPIN1 (phot1), also binds a flavin mononucleotide that acts as a chromophore.
PHOT1 protein has been localized to the plasma membrane in cells of dark-adapted hypocotyl tissue. Upon exposure to blue light, PHOT1 undergoes autophosphorylation and is relocalized to the cytoplasm. One role of PHOT1 appears to involve interacting with the auxin efflux carrier ABCB19 and causing its inhibition through phosphorylation. When ABCB19 is inhibited, auxin accumulates in the tip of the stem where it undergoes lateral transport to the shaded side of the stem through an efflux carrier.
After germination, plant growth is characterized by a the urgent need for the seedling to reach the light. During this period of growth, the seedling is throwing all of its resources into finding light. This kind of development, known as skotomorphogenesis, is characterized foremost by rapid elongation of the shoot, which is coupled with minimal elongation or further development of the root system. The shoot elongation that occurs during this period is strongly gravitropic, as this is one of the few cues available to the young seedling for orienting its growth. In dicot species, the energy-rich cotyledons are protected throughout this process of rapid stem elongation by the formation of an apical hook that tucks the cotyledons downward. Finally, dark-driven development is characterized by the lack of pigment biosynthesis, due in large part to the requirement for light as a catalyst in the production of chlorophyll. The seedling that develops in the dark is said to be etiolated.
Upon exposure to light for the first time, a number of changes occur to the seedling. The most obvious change is in the rate of stem elongation, which immediately slows and gives way to radial expansion. This is a very rapid response, occuring within seconds of the seedling sensing blue light. Almost concurrently with this, the primary root begins to elongate and will go on to begin producing lateral roots over the ensuing days. No longer needed for protection, the apical hook straightens and the cotyledons begin expanding and becoming green due to the synthesis of chlorophyll and other pigments required for photosynthesis.
The activation of such dramatic change in the transition from dark- to light-grown development requires many factors including several photoreceptors and many downstream signaling components. The rapid inhibition of stem elongation is mediated by a blue-light photoreceptor called cry1 (for cryptochrome). Many of the other responses associated with light are mediated by the phytochromes.
To identify other signal components of the photomorphogenesis pathway, mutant screens have been employed. One such screen was carried out to identify mutants that developed as though they had been exposed to light even though they hadn’t. These mutants were designated the constitutive photomorphogenesis (cop) mutants. The cop1 mutant was shown to have a lesion in a gene encoding an E3 ubiquitin ligase that targets transcription factors for degradation. These transcription factors regulate light-dependent genes, and by targeting them for degradation, COP1 acts as a negative regulator of photomorphogenesis. Many of the other COP genes encode components of a signalosome involved in regulating the localization of COP1 to either the nucleus or cytoplasm.
Following gravity perception, the sedimentation of amyloplasts must be converted into cellular information. Foremost among the candidates thought to specify this information is the formation of an auxin gradient.
Several different experimental approaches have shown that an auxin gradient does form during gravitropism. Using a radiolabelled-IAA approach, many researchers have confirmed that an auxin gradient forms throughout the elongation zone of gravity-stimulated roots. One of the limitations of this approach is the need to isolate a significant quantity of tissue, making it difficult to correlate the formation of the auxin gradient within the cells thought to be sensing gravity at the cap. More recently, an auxin gradient has been detected across the root cap columella cells by using an auxin-responsive promoter fused to GFP. This approach allows for the visualization of the effects of auxin within the cell, namely the resulting expression of a gene having a synthetic auxin response element. Because the GFP reporter protein must be produced and folded, there is a lag of approximately 1.5 h before a gradient of GFP appears following gravistimulation.
Another line of evidence that links gravity sensing with auxin redistribution is the very rapid relocalization that occurs to PIN3 proteins following gravistimulation. In vertical roots, PIN3 is localized uniformly throughout the plasma membrane. Within minutes after reorientation, PIN3 becomes localized to the new lower face of the cell, where it presumably directs auxin flux toward the lower flank of the root and inhibits their elongation. This relocalization is not due to the production of new PIN3 proteins, but rather results from the re-uptake of PIN3 from the plasma membrane via endocytosis. This is followed by the targeted exocytosis of vesicles containing PIN3 to a specified face of the plasma membrane.
While both of these lines of evidence support a link between gravity perception and auxin transport, the connection between the sedimentation of amyloplasts and redirection of PIN proteins remains loose. By crossing the auxin-responsive GFP into starchless mutants, we have shown that starchless roots fail to produce an auxin gradient across the root cap. This further supports the link between statolith sedimentation and the control of auxin transport.
In addition to auxin, several other potential growth regulating signals have been implicated in gravity signaling. For example, one of the fastest changes to be observed in gravity sensing cells is a change in ionic currents, measured as both membrane potential changes and surface potential changes. These changes have been reported within seconds after gravistimulation. Similarly, changes in pH have also been measured within minutes of gravistimulation, with root cap columella cells becoming alkaline and the apoplasts of these cells becoming acidic as a result of proton pumping. As of now, it is unclear whether or how these signals carry information about polarity that influences growth.
When a plant organ detects, through starch statolith sedimentation or another means, that it is no longer oriented in its preferred direction relative to gravity, a series of cellular and molecular events is initiated that results in a change in its direction of growth. This response, known as differential growth, leads to the eventual curvature of the organ back to its preferred orientation.
Some of the earliest studies of differential growth regulation focused not on gravitropic stimulation, but rather on phototropic stimulation. Charles Darwin was among the first plant scientists to study this phenomenon carefully:
…when seedlings are freely exposed to unilateral light, some influence is transmitted from the upper to the lower part, causing the latter to bend…
Charles Darwin, The Power of Movement in Plants, 1881
This summary came after careful experiments in which he shaded (or painted!) the tips of the growing seedlings and concluded that, while light seemed to be mostly perceived near the growing tip, the response occurred farther down the stem. Thus was set off a search for the nature of the “influence” that was transmitted, eventually leading to the discovery of the plant hormone auxin.
In a series of follow-up experiments to Darwin’s, it was shown that the factor causing differential growth could be extracted from rapidly growing tissues by placing tissue segments on a block of gelatin. The gelatin could then be used as a source of growth factor, and differential growth could be induced by applying the gelatin to one side of intact seedlings. These experiments suggested that differential growth begins with an unequal distribution of auxin, a concept known as the Cholodny-Went theory, first proposed in 1927.
A significant collection of physiological and biochemical evidence accumulated in support of the Cholodny-Went theory, including the discovery that auxin is transported in a polar manner through many organs of the plant. These data pointed to the need for a cell to have a mechanism for controlling the direction of auxin transport. This mechanism has now been identified to consist of several different kinds of auxin transporters, including the efflux carriers (PINs and ABCBs) and influx carriers (AUX1 and LAX3).
Immediately following germination, as the root emerges from the seed, it is faced with a decision: which way to grow? From the standpoint of adaptation, roots need to find a source of water and minerals quickly in order to sustain growth, both their own and that of the emerging shoot. Because water always flows downhill, the question of which way to grow seems to be reducible to, “Which way is down?”
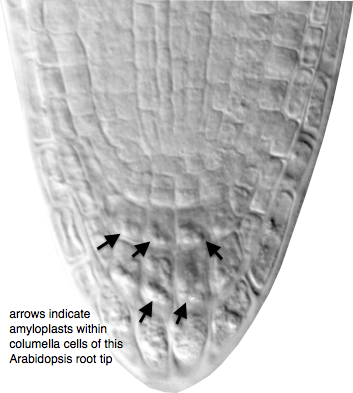
In order for the plant to detect its position in a gravity field, it must have some way to transduce the acceleration force due to gravity into cellular information. Most evidence points to the change in position of starch-filled plastids (called amyloplasts) within the cell as playing a central role in sensing the position of the organ in the gravity field. As an organ elongates and becomes displaced from vertical, these amyloplasts sediment to the new lower side of the cell due to their high density relative to the cytoplasm. These amyloplasts, therefore, act as statoliths, and their role in sensing gravity is known as the starch-statolith hypothesis.
Root tips of wild type and starchless mutants showing the presence or absence of starch statoliths in the columella cells of the cap.
There are several strong pieces of evidence to support the starch-statolith hypothesis. Plants that are unable to synthesize starch due to a mutation in a gene encoding a key enzyme in that process show a significant reduction in gravity responses, even though their growth rates are nearly wild-type. Similarly, mutants with intermediate levels of starch biosynthesis show gravity responses that correlate with the amount of statolith movement. Thus, full gravity sensitivity seems to require a full complement of starch within the amyloplasts to provide the density necessary for statolith sedimentation.
If the movement of statoliths within the gravity-sensing cells is sufficient to induce a growth response, then causing the movement of the statoliths by some other means besides gravity stimulation should induce a growth response as well. In a clever series of experiments, one team of researchers accomplished just such displacement of statoliths in vertically-growing roots. By taking advantage of the diamagnetic properties of starch, coupled with the ability to displace a diamagnetic material in a high-gradient magnetic field (HGMF), they induced root curvature similar to gravitropic growth by ‘pushing’ the statoliths within the cell.
A final kind of evidence implicating statolith sedimentation in gravity sensing comes from experiments in which the tissue containing the statoliths is somehow disturbed. The earliest reports of this kind of disturbance date to the mid-19th century. In roots of some species, the root cap is relatively easy to surgically remove without damaging the meristem. Upon removal, the root loses its ability to respond to gravity, and it slowly regains it as the cap is replaced through continued cell divisions. In a much more modern interpretation of this decapping experiment, investigators used laser ablation to obliterate specific tiers of the columella cells in the root cap, concluding that much of the gravity sensing appears to reside in cells in the second and third tiers of the columella.
Unlike in the root, where much gravity sensing appears to be localized to the root cap columella cells, sensing is distributed throughout the growing region of the young shoot. In dicots, sedimenting amyloplasts are found in the endodermis, a cyclinder of tissue throughout the stem just outside the vascular cylinder. In monocots, which produce a protective sheath called a coleoptile that encloses the emerging leaf, amyloplasts are distributed throughout the sheath.

 Immediately following germination, as the root emerges from the seed, it is faced with a decision: which way to grow? From the standpoint of adaptation, roots need to find a source of water and minerals quickly in order to sustain growth, both their own and that of the emerging shoot. Because water always flows downhill, the question of which way to grow seems to be reducible to, “Which way is down?”
Immediately following germination, as the root emerges from the seed, it is faced with a decision: which way to grow? From the standpoint of adaptation, roots need to find a source of water and minerals quickly in order to sustain growth, both their own and that of the emerging shoot. Because water always flows downhill, the question of which way to grow seems to be reducible to, “Which way is down?”
